
抽象的
背景
植物藻毒素样半胱氨酸蛋白酶(PLCPS.)是一类蛋白质水解酶,在根瘤共生(RNS)中起着重要作用PLCP公司豆类中的家庭基因非常有限,以及角色Glycine Max Plcps.(gmplcps.)在结瘤过程中,对结节的发育和衰老还没有完全了解。
结果
在本研究中,我们确定了97gmplcps.并进行了一个基因组的调查来探索大豆的扩张PLCP公司家族基因及其与RN的关系。19个亚拉加罗对基因组段,由77组成gmplcps.通过全基因组重复(WGD)事件形成的,鉴定出现,显示重复的高度复杂性。不同物种之间的系统发育分析表明谱系分化gmplcps.发生在家系扩展后,大串联重复片段在大豆中具有特异性。表达模式gmplcps.在共生相关的组织和结节中鉴定了相关的RNS相关gmplcps.并提供了在大豆中投入其推定的共生功能的见解。共生函数分析表明RNS相关GMPLCP.基因(glyma.04g190700)真的参与旋结和结节发展。
结论
我们的发现提高了我们对豆科植物功能多样性的理解PLCP公司家族基因,并为豆类的推定作用提供了见解PLCPS.在结瘤过程中,根瘤发育和衰老。
背景
植物藻素状半胱氨酸蛋白酶(PLCPS)是一种大类与植物病原体/害虫相互作用,种子萌发,发育,免疫,衰老,环化和应力反应相关的大类蛋白水解酶[1.,2.,3.,4.,5.,6.,7.,8.].PLCP属于C1A CYSPROT(家族C1,CLAN CA),是植物半胱氨酸蛋白酶的典型成员[3.,9].这些酶作为惰性前体制备,用信号肽,自动抑制前驱蛋白和活性蛋白酶域[1.].此外,一些plcp在其c端区域携带GRAN域[10.].PLCP公司家族基因已被系统地研究拟南芥,橡胶,木薯,蓖麻,杨树,葡萄树,gossypium hirsutum,卡里卡番木瓜和米[1.,11.,12.,13.,14.],虽然研究PLCP公司家族基因在豆类的整个基因组中非常有限。
PLCP公司家族基因已被证明参与结瘤[15.,16.]以及大豆的结节发育和衰老,黄芪西科斯,Pisum sativum.和截形苜蓿[17.]. 例如,PsCYP1在不确定结节中的衰老开始表达,PsCyp15A型和mscyp15a.在结节中表达[18.,19.].MTCP6.在发育和应激诱导的结节衰老中诱导表达,其早期表达促进结节衰老[20.].MTCP77通过加速植物PCD和ROS累积来确定结节衰老[21.].Asnodf32.,结节特异性半胱氨酸蛋白酶[22.]负面调节结节发育,组织衰老和结节寿命A. SINICUS.[23.].大豆CYSP1(gmcysp1.)可能参与结节发育和衰老[24.].然而,这些研究主要基于个别成员PLCPS..在大豆,数十个PLCPS.与根结节共生(RNS)有关[25.,26.,27.],虽然的作用GMPLCP.在旋转瘤中,不完全理解结节发育和衰老。
在本研究中,进行了全基因组调查,探讨了大豆的特殊特点和扩展PLCP公司家庭基因。表达概况gmplcps.在大豆根结节中分析共生以识别RNS相关的PLCPS.. 共生功能分析表明,一个RNS与GMPLCP.基因(glyma.04g190700)可能在结瘤和结节发育中发挥作用。我们的发现提高了我们对豆科植物功能多样性的理解PLCP公司家族基因,并为豆类的推定作用提供见解PLCPS.在结瘤过程中,根瘤发育和衰老。
结果
鉴定PLCP公司在大豆的基因家庭
大豆基因组的调查初步鉴定了106个基因基因座编码推定的PLCP甘氨酸最大var。威廉姆斯82基因组(表S1.)。所鉴定的大豆PLCP具有各种分子量,其各种分子量为6128.82-57,479.13Da,它们编码了具有55〜517个氨基酸残基和等电点(PI)的肽4.3〜9.32。NIH / NLM / NCBI CD搜索工具分析了这些106 PLCP中的保守域,以及表S2.列出详细信息。抑制剂_I29和肽酶_C1基序在大豆PLCPs中普遍保守,5个PLCPs(Glyma.04G028300、Glyma.10G120700、Glyma.13G229100、Glyma.14G085800和Glyma.17G239000)也有GRAN基序。此外,在这106个大豆PLCP中,有9个根据其序列长度被认为是假定的假基因(表S)1.)以及肽酶C1结构域缺失或肽酶C1结构域中存在大片段缺失(表S)2.),所以我们在大豆中识别出97个GMPLCP。
染色体地点和大豆重复PLCPS.
研究大豆的潜在重复性PLCPS.首先,候选人GMPLCP.位于一对副骨块中的重复对甘氨酸WGD。如图1所示。1.,含有77豆豆的19个候选副潜级段对PLCPS.在17豆蔻染色体上观察到。此外,两个基因簇(Glyma.06G275100,glyma.06g275200.和格莱玛06G275300;glyma.06g283000.和glyma.06g283100在染色体6上观察到)和glyma.12g130300.在染色体12上的大基因簇中定位。第二,在候选重复段对中进一步进行了共同性分析。2.示出了候选重复段对的侧翼区域。最后,由19个重复的段对甘氨酸确定了WGD事件(表1.). 值得注意的是,第10对和第11对在6号染色体或12号染色体上有较大的串联重复片段,表明这些大豆中存在高度的复杂性PLCP公司复制品。

染色体位置,串联区域复制和19对旁级段gmplcps..红色虚线表示重复的段对。由大豆最近WGD事件产生的重复段由不同的彩色框表示。染色体数目在每条染色体上方表明gmplcps.不在CHR01中。地图图表和Adobe Illustrator分别用于生成和修改图形

19级副基因段对的相机性分析gmplcps..复制GMPLCP.基因对由绿色和紫色箭头或棍子表示。基因在DNA正链(向左)和负链(向右)中的位置由箭头方向指示
利用大豆基因组数据库检索这些基因的同义突变率(Ks)值PLCPS..所有KS值范围为0.1028至0.3374,而这19个重复的副吡咯的分歧时间为8.43和27.66 mya,平均为14.2 mya(表1.),这与WGD事件(10–20 Mya)一致。非同义突变率(Ka)与Ks(ω)之比 = Ka/Ks)通常用于测量作用于编码序列的选择历史[28.].当ω<1时,至少一个基因在净化选择下,而ω> 1表示定向选择[29.,30.].表格1.表明,对于17个重复的段对(第10号和第11号除外,ω为0.069至0.643,表明这些基因受净化选择受到限制。
系统发育和外显子性结构分析PLCPS.在拟南芥蒂利亚纳,M.血液凋亡,莲花japonicus.和大豆
探讨大豆的系统发育关系PLCPS.随着其他豆科植物和非豆科植物的那些,我们在97中进行了全长的肽序列对齐gmplcps.(大豆),26atplcps.(A. Thaliana.), 33mtplcps.(M.血液凋亡)和25岁ljplcps.(L. japonicus.)使用MEGA(6.0版本)(图。3.)。这个PLCPS.从这四种不同的物种分布在九个基团中(A组给A组)。其中,大豆PLCPS.和19个重复的分段对甘氨酸WGD事件分布在所有九个群体中,表明大豆的谱系分化PLCPS.发生在家庭扩张之后。大豆PLCP公司在这9个类群中,成员分布不均匀,其中A、B、D组仅以豆科植物组成PLCPS.(图。3.),说明这三组在分化前就发生了A. Thaliana.和豆科植物或是A. Thaliana.或特定于豆类。除了,AT5G45890是独立于每组,表明该基因具有特异性特征和新功能。

系统发育关系PLCPS.来自大豆,L. japonicus.,M.血液凋亡和A. Thaliana..程序Mega6.0用于构建包含97的系统发育树PLCPS.来自大豆,25岁PLCPS.从L. japonicus.,33.PLCPS.从M.血液凋亡和26PLCPS.从A. Thaliana.. 这些PLCPS.被分为九个群体,即A组I.基因复制事件标有“ ”
”
之间PLCPS.虽然在三种豆科植物中发现了串联重复事件M.血液凋亡和L. japonicus,大串联重复片段对只存在于大豆中PLCPS.(对A组和B组的10和11),表明这种复杂的复制PLCPS.是针对大豆,而不是豆类。数量PLCPS.在大豆中显着超过了M.血液凋亡和L. japonicus.除上述大串联重复片段事件外,其主要原因可能是大豆中的片段重复事件,特别是G组和I组(图1)。3.)。在这两组中,只有三个mtplcps.和三ljplcps.,虽然有20个副吡咯大豆PLCPS.. 然而,这些同源基因在大豆中保留的原因尚不清楚,这可能与大豆的某些生物学功能有关。
每个基因的cDNA序列PLCP公司在上述九个基团中(图。3.)与其基因组序列进行比较,以分析它们的UTR /外显子结构,并且在每组中发现了类似的基因结构PLCPS.(图。4.)。大部分的PLCPS.在A组到D组D表现出相对简单的基因结构,其中70%PLCPS.只包含一个内含子。E组到I组,所有PLCPS.在这五组中有两个或更多的内含子(图。4.). 这些群体特异性基因结构与它们之间的关系是一致的PLCPS.跨物种并进一步支持这些功能分歧PLCPS..

外显子/内含子结构PLCPS.来自大豆,L. japonicus.,M.血液凋亡和A. Thaliana..181.PLCPS.被分为九个组,即A组到A组(根据图1中的分类。2.)。淡绿色盒代表外显子,李子红色代表UTR,黑线是指内含子,斜杠代表过长的内含子,顶部的鳞片用于估计外显子和内含子的尺寸
对大豆的分析PLCPS.
去大豆PLCPS.基于使用大豆基因组数据库中的数据的42个GO术语调查。这些GO功能术语分为三类:生物过程,细胞组分和分子功能(图。5.),它们的详细基因ID信息显示在表S中3.. 其中,5个基因(glyma.06g042600.,glyma.06g282300.,glyma.12g124300.,格莱玛12G126000和glyma.12g131000)没有预期的Go功能。其余均具有半胱氨酸型肽酶活性,并参与了蛋白质的水解过程,其中大部分大豆为半胱氨酸型肽酶PLCPS.均定位于细胞外区(87个基因)。对于重复片段对,在对1中有两个基因,在对15中有三个基因,在最复杂的两对中有23个基因(对NO。10和11)参与衰老、乙烯反应、真菌防御反应、不相容互作和叶片衰老过程,并具有衰老相关液泡位置。在1、9、17和19对基因中,其中一个串联重复序列基因具有不同的功能。另外,其余12对基因对中,除第6对(glyma.06g042600.没有预测的去功能),两种基因都参与了相同的生物过程。

基于本体本体论的功能诠释gmplcps.. 三个GO域-分子功能(1-4),生物过程(5-31),细胞成分(32-42)如图所示。每个术语中的基因数用直方图表示
大豆的表达模式PLCPS.在RNS.
确定大豆不同成员之间的系统发育关系PLCPS.,我们基于97全长PLCP肽序列比对进行系统发育分析。结合图1中的分类。2.,这97豆豆PLCPS.被分成9级,例如大豆PLCPS.A组分为1类,大豆PLCP公司B组中的S分为2等级,等等(图。6.一种)。

已鉴定的97个基因的系统发育和表达分析gmplcps..A.鉴定97的系统发育分析gmplcps..系统发育树建设gmplcps.利用RAxML、MEGA版本6.0和MrBayes 3.2等程序对氨基酸序列进行了全长推导。该树显示了9个主要的系统发育类群(类群1到类群9),用不同的阴影表示。B已鉴定的97基因的表达分析gmplcps.在植物血统数据库中的五个相关组织中。表达值gmplcps.在Phytozome数据库中,R中的Pheatmap包用于产品热图。这些组织包括结节(1),根毛(2),根(3),结节。共生。条件(4)和根。共生。条件(5)。C对鉴定出的97gmplcps.在五种不同的大豆根瘤中。我们之前的RNA序列数据[18.[R中的Pheatmap封装用于产品的热图。这五个结节样品包括分支阶段(6),开花阶段(7),结果阶段(8),Pod阶段(9)和收获阶段(10)中的结节,并显示了这五个重要的发展阶段的描述我们以前的研究[18.]
确定哪种大豆PLCPS.参与RNS,我们首先调查了97豆豆的表达概况PLCPS.在植物共生相关组织数据库的基础上,建立了植物共生相关组织数据库,这些组织包括根、根毛、根瘤、根瘤。共生条件与根系。符号状态。结果表明,大豆PLCPS.在这些组织中具有不同的表达模式,并且大多数高表达基因主要关注在6级至10级(图。6.b)。然后,我们使用我们以前的RNA-SEQ数据[18.]分析了这97种大豆的表达谱PLCPS.根据我们之前的RNA序列数据(图。6.C)。我们比较了这97豆豆的表达水平PLCPS.在上述与共生相关的组织和结节样品中,并在与共生相关的组织中高表达的基因中,有些PLCPS.在结节样品中也具有高表达(图。6.b和c),表明这些PLCPS.可以参与RNS或结核发育。
考试是否这些PLCPS.在共生相关的组织和结节样品中的高表达具有结节发育和/或衰老的作用,我们分析了28所选择的表达差异PLCPS.通过QPCR不同的结节开发时间点之间。首先,四种参考基因的表达稳定性(elf1b.,Qact.,G6PD公司和泛素),其中,elf1b.和Qact.所有样品中最稳定,而且GMG6PD.和泛素始终不稳定(图S1.)。然后,elf1b.和Qact.结果表明,这28个PLCPS.在五个结节之间有差异表达(图。7.)。其中,12PLCPS.在结节发育和/或衰老期间上调,并在接种后84天收集的结节处达到它们的峰(图84dN)(图。7.a、 d、g、l、n、r、s、u、v、x、y和ab)。四PLCPS.在结节发育过程中表达下调,在64dN或84dN时表达较低(图。7.B,C,F和H)。七PLCPS.在30dn或42dn达到峰值(图。7.e、 I、k、m、o、w和aa)。三PLCPS.在结节发育和/或衰老期间被下调在调节上调(图。7.p,q和z)。glyma.06g014800.在结节发育和/或衰老期间被上调然后下调(图。7.j),和格莱玛14G216300在30dn和84dn处具有高表达(图。7.t) 是的。

28例患者qPCR分析gmplcps.结节发展。大豆elf1b.和Qact.作为参考基因,使每个基因的相对表达水平正常化GMPLCP.基因,12dN中的表达水平用作起始点控制,以计算其他结节样品的相对表达水平。重复所有QPCR反应三次,数据显示为平均值±SD
功能分析glyma.04g190700大豆毛状根转化的研究进展
如上所述,glyma.04g190700在共生相关的组织中高度表达(图。6.b) 并在结节发育过程中上调(图。6.c和图。7.g),建议glyma.04g190700可能在结瘤和结节发育中起作用。为了证实这一结果,RNA干扰glyma.04g190700采用大豆毛状根转化法(图。8.)。与BXYD3接种后50天的共生表型在50天内得分.rnaiglyma.04g190700导致结节数增加(图。6.一种)。RNAi的结节glyma.04g190700显示比对照显着更高的氮酶活性(图。6.b) 是的。表达水平glyma.04g190700和四种结果蛋白基因(ENOD40,结节35,钙调蛋白和LB1.)[31.,32.,33.通过毛发根和结节的QPCR检查(图。8.C和D)。这个glyma.04g190700与对照组相比,RNAi毛状根和根瘤中的转录物减少到50%以下(图。8.C和D)。四种结核蛋白基因的表达水平(ENOD40,结节35,钙调蛋白和LB1.)急剧增加了glyma.04g190700与控制毛根和结节中的那些相比,RNAi毛的根和结节(图。8.综上所述,这些结果强烈表明glyma.04g190700参与结核和结节发展。

效果glyma.04g190700在大豆的RNS上的RNAi。A.每个植物的平均点数有变化glyma.04g190700转录水平。每个样品中的独立转基因植物的数量在括号中表示。B对照中结节的氮酶活性和glyma.04g190700RNAi植物。以p5941-35S内含子(GFP-Bar标记)转基因毛状根为对照(A.和B)。CQPCR分析转录水平glyma.04g190700,ENOD40,结节35,钙调蛋白和LB1.在对照和转基因长毛根中。DQPCR分析转录水平glyma.04g190700,ENOD40,结节35,钙调蛋白和LB1.在对照和转基因结节中。从根系系统(包括小结节)中分离的总混合RNA或多个独立转基因植物的结节用于QPCR分析。控制毛状根和结节中的参考用于计算转基因毛状根和结节中这五种基因的相对表达水平。数据显示为平均值±SD和“ “表示不同接种毛根之间的统计差异(T测试,P < 0.01)
“表示不同接种毛根之间的统计差异(T测试,P < 0.01)
讨论
PLCPS.是一大类蛋白水解酶,在RNS中发挥重要作用[7.,8.],虽然全基因组研究PLCP公司豆科植物的家族基因非常有限。本研究首次对大豆进行了全基因组调查PLCP公司基因并探索大豆的扩张PLCP公司家族基因。结果的表达模式gmplcps.在大豆中,RNS为豆科植物的假定作用提供了见解PLCPS.在结瘤过程中,根瘤发育和衰老。
基因组扫描和大豆的扩展PLCP公司家族基因
基因组识别PLCPS.已系统地执行拟南芥,橡胶,木薯,蓖麻,杨树,葡萄树,G. Hirsutum,C. Papaya.和米[1.,11.,12.,13.,14.].在本研究中,97gmplcps.在大豆中鉴定出,且数量多于以往研究中的其它植物。根据系统发育分支和结构特征,这些大豆PLCPS.被分为九个亚科,这与以前的研究相似[11.,34.,35.,36.].然而,在这九个群体中,只用豆类形成三组PLCPS.而且只有26岁在PLCPS.在群体中,与其他研究有很大差异[1.,11.,12.].
两种全基因组重复(WGD)事件发生约59和13 mA,在大豆基因组中经历了[37.,38.,39.],对大豆中许多多岛家族的扩展具有显着贡献[28.,40,41.].不同的基因家族可能有不同的多倍性衍生的重复事件和重复基因保留的进化机制[28.],它在家庭的自适应演化和生物学功能中起重要作用[42.,43.,44.].大豆PLCP公司基因家族中,数量明显多于A. Thaliana.,M.血液凋亡和L. japonicus.,这可能是由WGD事件,串联复制和大规模的分段复制来引起的[11.,12.].在大豆的重复事件中PLCP公司基因家族两个大型节段复制对包括37gmplcps.小串联重复对和单基因片段重复对大豆的扩展也有贡献PLCP公司家庭基因。此外,这些重复簇基因的保留可能主要归因于它们不同的表达模式,在这些对中的特殊结构和功能,这与先前的研究相似[45.,46.,47.].
潜在的共生功能gmplcps.在大豆
以前的研究表明PLCPS.在内源蛋白质转换中发挥重要作用[48.]种子特征,萌发和成熟[49.,50.非生物环境压力[11.,49.]及保护植物免受螨害[51.], 菌类 [52.,53.], 细菌 [54.[病毒[55.].根瘤菌III型分泌系统(T3S)是植物病原体的毒力因子的导引子,可以通过豆类衍生的类黄酮诱导,并据报道通过主体防御系统识别来调节染色过程[56.].此外,我们以前的研究表明,结节发育和衰老与植物免疫防御直接相关[26.].PLCPS.据报道涉及生物和非生物应激[11.]负责对病原体细菌进行防御,并调节植物免疫力[54.]近年来发现了越来越多的分子机制[11.].在本研究中,值得注意的是,没有大的串联复制对,只有两个小串联复制对gmplcps.在结节发展中表达了脱节,表明RNS相关GMPLCP.重复基因主要来源于单基因片段重复,而不是串联重复。与以往研究不同的是,RNS相关重复对中的两个同源基因在RNS中表现出相似的表达模式[11.].
已经提出了以前的研究PLCPS.在几种豆科植物根瘤的发育和衰老中起重要作用[17.,20.,21.,22.,23.,24.,57.].在大豆,PLCPS.可能在结瘤中起重要作用[15.,27.]以及结节发育和/或衰老[25.,26.].然而,角色的gmplcps.在结瘤中,结瘤的发育和衰老尚未完全了解。在目前的研究中,表达谱gmplcps.在五种共生相关的组织和五种不同的结节样本中[18.]全面分析,结果确定了数十个相关的PLCPS.,建议多个gmplcps.可以参与结节信号识别和免疫和/或结节发育和衰老。在以前的转录简介数据中,表达式为14gmplcps.在衰老发作期间特别增加[25.].在这些当中gmplcps.,在结节衰老期间,12个基因上调,而其余两个基因在64dN或84dn下具有低表达。在这项研究中,其他四个gmplcps.也可能在结节发育和衰老中发挥作用。此外,五PLCPS.可以参与氮固定过程,八个PLCPS.可以参与结核发育的早期和/或中期。这些数据表示gmplcps.可能不仅参与结瘤衰老,而且参与结瘤和结瘤发育。在本研究中,共生性功能分析glyma.04g190700表明它真的参与了染色结和结节发育。RNS相关的具体监管角色和不同功能gmplcps.需要进一步研究。
RNS相关的特征gmplcps.
以前的研究表明,多倍体衍生的重复基因导致豆类中的增强RN [58.,59.].在本研究中,RNS相关gmplcps.主要集中在第5类到第9类,这些基因大部分参与了单基因片段的复制,说明多倍体起源于基因的重复事件gmplcps.在RNS中也发挥了重要作用。此外,RNS相关gmplcps.具有含有UTR序列和三个或更多内含子的相对复杂的基因结构。在这些RNS相关的GMPLCP中观察到三种发散的图案模式。第一个模式包含了Propeptide_c1域[60.]肽酶_c1a_cathepsin b域[61.](仅适用于Glyma.03G226300和Glyma.19G223300)。在三种蛋白质中发现的第二种模式(Glyma.04G028300,Glyma.14G085800和Glyma.17G239000),不仅具有两个典型的基序(抑制剂_I29结构域和肽酶_C1结构域)[62.[还有一个Gran畴,也是与一些已知的plcps相似[10.].其余的或大多数相关的RNS相关的GMPLCP通常被分类为包含两个典型图案的第三种模式。这些结果表明RNS相关GMPLCP中没有特殊的蛋白质结构,类似于染色和结节显影相关的大豆胱抑素[63.].
结论
总之,我们进行了一个基因组调查并确定了97gmplcps..对由WGD事件产生的19个片段重复对进行了鉴定和分析,表明大豆的重复具有高度的复杂性PLCPS..表达概况gmplcps.利用大豆根瘤共生体鉴定了RNS相关基因PLCPS..共生函数分析表明RNS相关GMPLCP.基因(glyma.04g190700)真的参与旋结和结节发展。我们的研究结果改善了对豆类功能多样性的理解PLCP公司家族基因,并为豆类的推定作用提供见解PLCPS.在结瘤过程中,根瘤发育和衰老。
方法
鉴定PLCPS.在大豆中,M. Truncatula.,L. japonicus.和A. Thaliana.基因结构分析
这个PLCP公司从大豆基因组数据库中鉴定了大豆家族基因[http://soybase.org/]以及甘氨酸最大wm82.a2.v1 phytozome数据库[http://www.phytozome.net/soybean.]. 用NIH/NLM/NCBI-CD搜索工具对所有已鉴定的GmPLCPs进行分析,分析其不含肽酶C1结构域或肽酶C1结构域存在大片段缺失和/或其序列长度< 150个氨基酸被认为是假定的假基因,并被手动删除(表S)1.和表S.2.). 采用基本的局部比对搜索工具算法(BLASTP),以e值<1e-10为阈值,进行同源序列的识别gmplcps.在M. Truncatula.,L. japonicus.和A. Thaliana..外显子/内含子/ UTR结构PLCPS.通过使用基因结构显示服务器程序GSDS2.0进行分析(http://gsds.cbi.pku.edu.cn/)。
大豆PLCPS.顺序
检索phytozomev12.0数据库下载gmplcps..expasy网站(http://web.expasy.org/protparam/)计算了其等电点和分子量gmplcps..地图图表软件和大豆基因组注释文件(GMAX_275_WM82.A2.v1.gene.gff3)用于分析gmplcps.在染色体上。大豆和大豆饲养员的工具箱(https://soybase.org/gb2/gbrowse/gmax2.0/)用于获得被视为近期重复的块。
系统发育分析
不同的PLCPS.应用于多种系统发育分析,包括97gmplcps.,26PLCPS.从A. Thaliana.,33.PLCPS.从M.血液凋亡,和25L. japonicus..共克尔W程序用于进行全长肽序列比对。Mega6软件[64.[邻居加工(NJ)方法和1000个引导复制与P距离模型的分析用于执行多种系统系统发育树。程序RAXML,MEGA V6.0和MRBAYES 3.2(http://www.megasoftware.net.)[64.,65.,66.]进行了97的系统发育分析gmplcps..Raxml 8.0.0 [67.用于执行最大似然(m1)分析,使用raxml中的扩展多数规则共识树标准(Auto Mre)进行了1000个引导测试,用于执行Rapid 1000引导复制分析,混合模型用于构建MRBAYES分析。
大豆WGD事件重复片段对的识别与分析
为了识别由大豆WGD事件形成的重复片段对,首先确定每个片段对的同义词(Ks)GMPLCP.或重复的基因对gmplcps.是从大豆和大豆饲养员的工具箱中鉴定出来的。其次,根据分布的gmplcps.在大豆染色体和它们的同义值,鉴定了11串联重复基因簇(图。1.). 第三,初步鉴定了两个大的同源簇(10号和11号对),每对中有两个大的串联重复基因簇;5个同源簇(1号、9号、15号、17号和19号对),每对中有一个串联重复基因簇;12个同源基因对。然后,为了检验这些初步确定的19个候选旁同源片段对,本研究采用了以下两个标准:1)将重复片段对分组在一起gmplcps.系统发育树(图。3.和图。6.a)和2)在共聚合分析中显示候选重复段对的侧翼区域(图。2.)。
发散时间(T)用T = Ks/(2) × 6.1 × 10- 9)×10- 6mya [68.]估计复制对的日期。此外,使用PAML程序的ON00方法计算ParaLog对的非同义(KA)[69.]在重复后调查达尔文的阳性达尔文选择。计算Ka至Ks(ω= Ka / ks)的比率以测量在编码序列上的选择历史[28.].
Go注释与基因表达分析
围棋诠释gmplcps.通过在大豆基因组数据库中使用“GO术语富集工具”进行[http://soybase.org/].大豆数据库(https://soybase.org/Goslimu v2/仪表板。php)来下载这些Go术语的详细基因信息。植物群落数据库(植物群落12,http://www.phytozome.net/大豆)用于下载表达式模式数据gmplcps.在五种与共生相关的组织中。表达模式gmplcps.通过检索我们以前的RNA-seq数据,对5个结节样本进行了分析[26.].r中的pheatmap包[41.用来产生这些的热量gmplcps..
植物材料和生长条件
在日光温室中,将表面灭菌的大豆天龙一号幼苗在湿润的滤纸上萌发,昼夜周期保持在16/8 h和相对湿度(RH)分别为70%和28% °C,持续2–3天。然后在盛有消毒珍珠岩和蛭石(比例为1:1)的盆中生长幼苗,并用半强度B&D培养基浇水[63.].4-5天后,使用大豆根茎113-2菌株(储存在我们的实验室)接种幼苗。从大豆结节中收集RNA分离的样品,在五个时间点:12dN(接种后12天结节),30dN(接种后30天结节),42dN(接种后42天结节),64dN(64天结节结节)在接种后)和84dn(接种后84天结节)。单独收集来自不同时间点的结节,用三种生物重复收集,并在-80℃下冷冻以进行RNA分离。
RNA提取和qPCR
我们使用TRIzol试剂(Invitrogen,USA)提取结节的总RNA,DNase I(Takara)消化总RNA,以及带有gDNA擦除器(Takara Bio,Inc.)的Prime Script RT试剂盒(完美实时)执行反向转录分析。使用Epoch多体积分光光度计系统、NanoDrop和Agilent 2100生物分析仪(Agilent Technologies,Palo Alto,CA,USA)测量RNA的数量和质量,并使用五个RNA样品上的qPCR反应来确认这些RNA样品中不存在gDNA。使用表S中列出的引物组,将来自约1μg RNA的逆转录的cDNA用作qPCR的模板4..RT-PCR扩增混合物(20μL)含有2μL模板cDNA,ITAQ血液SYBR绿色超混(10μL)(Applied Biosystems)和0.5μl前进和反向引物。在CFX连接实时系统(应用生物系统)上运行反应,每个测定包括无模板控制(负控制)。30s在95℃下的循环条件为30s,然后在95℃,在60℃下在62℃和12 s处在72℃下在72℃下的40℃和12℃下循环。PCR扩增后,为每个PCR产物产生熔化曲线以检查PCR反应的特异性(图S2.)。四种参考基因的表达稳定性(elf1b.,Qact.,G6PD公司和泛素)被评估,和elf1b.和Qact.被选为28所选择的QPCR分析的参考基因gmplcps.. 以两个参考基因为对照,采用geNorm方法对每个模板的样本周期阈值(CT)进行标准化[70]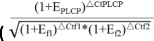 ;E, PCR效率)分析qPCR实验中基因表达的相对变化。每个样本使用三个生物复制样本和三个或三个以上的技术复制反应,以确保统计的可信度。
;E, PCR效率)分析qPCR实验中基因表达的相对变化。每个样本使用三个生物复制样本和三个或三个以上的技术复制反应,以确保统计的可信度。
Glyma.04g190700特定的RNAi
206bp的5′- 解释glyma.04g190700通过PCR扩增并克隆到P5941-35S - 内含子 - (GFP-BAR标记)中,产生pglyma.04g190700-Rnai-1;185磅的3'-区域的片段glyma.04g190700通过PCR扩增并克隆到P5941-35S - 内含子 - (GFP-BAR标记)中,产生pglyma.04g190700-rnai-2。在这两者中glyma.04g19070特定于特定的RNAi载体,感觉和反义glyma.04g190700RNA序列将与内含子分开的串联连接。用于构建这两个的底漆glyma.04g190700-特定的RNAi载体列于表S中5..
A. rhizogenes.细胞K599覆盆子glyma.04g190700-rna干扰-1,pglyma.04g190700-RNAI-2和空载体用于诱导大豆中转基因毛状根的形成。表达空载体的转基因毛状根用作对照。接种用BXYD3后,生长具有阳性转基因毛状根的植物50天,并评分染色表型。表达水平glyma.04g190700,ENOD40,结节35,钙调蛋白和LB1.在p中glyma.04g190700通过使用表S中列出的引物通过QPCR测定-RNAI或对照5.以及前面描述的过程[71.,72.].如通过气相色谱(GC-14,日本)所述的乙炔还原测定(ARA)测定氮酰酶活性[73.].
大豆毛状根转化
glyma.04g190700特定的RNAi构建体被转移到A. rhizogenes.通过电穿孔,然后A. rhizogenes.- 导明的程序[74.]诱导大豆毛状根形成。侵染后,将大豆幼苗移栽到含大豆全氮营养液的水培上,并用透明塑料覆盖以保证高湿度。愈伤组织形成后(约5 将幼苗移栽到大型水培池中。在两周内,毛状根开始从感染部位发芽,并用荧光显微镜进行筛选(图3.). 从转基因毛状根中提取基因组DNA,通过基因特异性引物证实CK和RNAi。每个幼苗只允许有一个转基因毛状根,并用绳子包裹。对于结瘤试验,转基因复合植物接种根序BXYD3并转移到含有1/10n(530mmol / Ln)的蛭石上的罐子,并在26°C的16- / 8小时/夜间循环中生长。我们在BXYD3接种后50天在50天内探测了这些转基因复合植物的结节表型,并使用了空载体转基因毛状根作为对照。
可用性数据和材料
本研究中使用的所有数据和材料可在合理的请求时从相应的作者获得。
缩写
- PLCP:
-
植物木瓜样半胱氨酸蛋白酶
- RNS:
-
根结节共生
- WGD:
-
全基因组重复
- 格兰:
-
肉芽桃
- PI:
-
等电点
- ks:
-
同义突变率
- K a:
-
非同义突变率
- RNAi:
-
RNA干扰
- NJ:
-
邻接
- ml:
-
最大似然
- RH:
-
相对湿度
- BLASTP:
-
基本的本地对齐搜索工具算法
- T3SS:
-
III型分泌系统
参考
- 1。
刘H,胡M,王Q,程L,张Z。木瓜蛋白酶样半胱氨酸蛋白酶在植物发育中的作用。前植物科学。2018;9:1717.
- 2。
Shindo T, Van der Hoorn RA。木瓜样半胱氨酸蛋白酶:植物及其入侵者在分子战场上的关键角色。Mol Plant Pathol. 2008;9(1): 119-25。
- 3.
木瓜样半胱氨酸蛋白酶在植物免疫中的作用。新植醇。2016;212(4):902 - 7。
- 4.
Díaz-Mendoza M,Velasco-Arroyo B,González-Melendi P,MartínezM,Díazie.C1A半胱氨酸蛋白酶 - 叶片衰老中的相互作用。J Exp Bot。2014; 65(14):3825-33。
- 5。
Grudkowska M,ZagdańskaB.植物半胱氨酸蛋白酶的多功能作用。Acta Biochim Po。2004; 51(3):609-24。
- 6。
Rabbani MA,Maruyama K,Abe H,Khan MA,Katsura K,Ito Y,等。利用cDNA微阵列和RNA凝胶印迹分析监测水稻基因在低温、干旱和高盐胁迫下的表达谱和脱落酸应用。植物生理学。2003;133(4):1755–67.
- 7。
Kempema La,Cui X,Holzer FM,Walling LL。Arabidopsis转录组的反应变化饲料饲喂Silverleaf粉虱若虫。对蚜虫的反应中的相似性和区别。植物理性。2007; 143(2):849-65。
- 8。
Rehm FBH, Jackson MA, De Geyter E, Yap K, Gilding EK, Durek T等。木瓜蛋白酶类半胱氨酸蛋白酶制备植物环肽前体用于环化。《美国国家科学院学报》上。2019; 116(16): 7831 - 6。
- 9。
罗林斯街。MEROPS数据库中的肽酶抑制剂。生物化学。2010;92(11):1463–83.
- 10。
贝特曼、贝内特•惠普。颗粒蛋白基因家族:从癌症到痴呆。BioEssays。2009; 31(11): 1245 - 54。
- 11.
张松,徐忠,孙浩,孙玲,沙班M,杨欣,等。陆地棉中木瓜样半胱氨酸蛋白酶的全基因组鉴定及对大黄萎病的功能表征。植物科学学报,2019;
- 12.
刘家,沙姆达A,Niewiara MJ,Singh R,Ming R,Yu Q.鱼豚霉素番木瓜:谱系特异性基因重复和扩张。BMC基因组学。2018; 19(1):26。
- 13。
王W,周XM,Xiong HX,Mao Wy,Zhao P,Sun MX。水稻木蛋白石和肉桂样蛋白酶:基因组鉴定,综合基因特征表征和表达分析。BMC植物BIOL。2018; 18(1):87。
- 14。
啤酒ep,琼斯am,dickerman aw。S8丝氨酸,C1A半胱氨酸和A1天冬氨酸蛋白酶家族拟南芥.植物化学。2004; 65(1):43-58。
- 15.
Quain MD,Makgopa Me,Cooper JW,kunert kj,voyer ch。异位植物植物表达式增加结节数并影响大豆的反应(甘氨酸最大)导致氮缺乏。植物化学。2015;112:179–87.
- 16。
Sheokand S,Brewin NJ。瘤瘤和氮固定中的半胱氨酸蛋白酶。印度j exp biol。2003; 41(10):1124-32。
- 17。
Mergaert P,Kereszt A,Kondorosi E.基因在氮固定共生结节细胞中的基因表达截形苜蓿以及其他结瘤植物。植物细胞。2020;32(1):42–68.
- 18。
Vincent JL, Brewin NJ。半胱氨酸蛋白酶在豌豆根瘤细胞空泡、囊泡和共生体中的免疫定位。植物杂志。2000;123(2):521 - 30。
- 19。
Kardailsky IV, Brewin NJ。半胱氨酸蛋白酶基因在豌豆根瘤发育和衰老中的表达。植物与微生物的分子相互作用。1996;9(8):689-95。
- 20。
Pierre O,Hopkins J,Combier M,Baldacci F,Engler G,Brouquisse R等。木瓜蛋白酶和肉桂蛋白酶参与衰老过程截形苜蓿结节。新植物。2014; 202(3):849-63。
- 21。
邓俊,朱福,刘继,赵y,文杰,王t等。转录因子BHLH2抑制半胱氨酸蛋白酶77以负面调节结节衰老。植物理性。2019; 181(4):1683-703。
- 22。
奈藤Y、富士M、Usami S、慕鲁卡Y、山田T。半胱氨酸蛋白酶在紫云英根瘤形成中的作用梅尔霍齐乌利普华县。rengei..植物理性。2000; 124(3):1087-96。
- 23。
李Y,周L,Li Y,Chen D,Tan X,Lei L等人。特异性植物半胱氨酸蛋白酶,Asnodf32.,参与结节衰老和绿色粪肥豆类黄芪石榴石的氮固定活性。新植物。2008; 180(1):185-92。
- 24。
Alesandrini F,Mathis R,Sype GV,Hérouartd,Puppo A.在大豆结节发育和衰老中的半胱氨酸蛋白酶和过氧化氢的可能作用。新植物。2003; 158(1):131-8。
- 25。
van wyk sg,du plessis m,cullis ca,kunert kj,vorster bj。半胱氨酸蛋白酶和胱抑素在大豆结节发育过程中的表达和活性和衰老。BMC植物BIOL。2014; 14:294。
- 26。
元股,李河,陈赫,张CJ,陈立,昊QN等。RNA-SEQ对大豆五种不同发育阶段的结节发育分析(甘氨酸最大)接种Bridyrhizobium japonicum菌株113-2.Sci Rep.2017;7:42248.
- 27。
袁S,李R,陈S,陈H,张C,陈L,等.大豆不同根瘤菌菌株差异基因表达的RNA-Seq分析(甘氨酸最大)根。前植物SCI。2016; 7:721。
- 28。
李士,王,姬d,薛z,yu y,江y等。大豆膜结合NAC转录因子基因的进化和功能分析。植物理性。2016; 172(3):1804-20。
- 29。
牧师N,Hoen Dr,Huynh Ml,Harrison PM,Bureau Te。大米宿主基因片段宿主基因段的进化命运。Genome Res。2005; 15(9):1292-7。
- 30.
弗里林M。亚功能化的进化位置被降级。基因组动力学。2008;4:25–40.
- 31。
Minami E,Kouchi H,Cohn Jr,Ogawa T,Stacey G.早期Nodulin,Enod40的表达,在大豆根中响应于各种脂肪丁蛋白信号分子。工厂J. 1996; 10(1):23-32。
- 32。
Legocki RP,Verma DP。来自大豆豆的结节特异性植物蛋白(Nodulin-35)。科学。1979; 205(4402):190-3。
- 33。
Choudhury SR,Pandey S.在大豆结节形成期间G蛋白循环依赖性调节。植物细胞。2015; 27(11):3260-76。
- 34。
Richau Kh,Kaschani F,Verdoes M,Pansuriya Tc,Niessen S,Stüberk,等。植物木质半胱氨酸蛋白酶的亚分类和生化分析显示亚家族特异性特征。植物理性。2012; 158(4):1583-99。
- 35。
马丁内斯M,迪亚兹I。植物半胱氨酸抑制素与其靶半胱氨酸蛋白酶的起源和进化表明了一种复杂的功能关系。BMC进化生物学。2008;8:198.
- 36。
Zou Z,谢克,杨L.木蛋白石样半胱氨酸蛋白酶编码橡胶基因(Hevea Brasiliensis.):比较基因组学,系统发育和转录分析分析。Planta。2017; 246(5):999-1018。
- 37。
Schlueter Ja,Dixon P,Granger C,Grant D,Clark L,Doyle JJ,等。采矿EST数据库解决主要作物物种中的进化事件。基因组。2004; 47(5):868-76。
- 38。
Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, Nelson W,等。古多倍体大豆基因组序列。自然。2010;463(7278):178 - 83。
- 39。
Vanneste K,Baele G,Maere S,Van de Peer Y. 41植物基因组的分析支持与白垩纪 - 古近边界相关的成功基因组重复浪潮。Genome Res。2014; 24(8):1334-47。
- 40。
刘HJ,唐ZX,汉XM,杨ZL,张FM,杨HL等。大豆GST超级家族中酶活性的分歧提供了新的洞察全基因组复制品的进化动态。mol Biol Evol。2015; 32(11):2844-59。
- 41。
韩y,li x,cheng l,liu y,wang h,ke d等。大豆JMJC域蛋白质的基因组分析表明全基因组重复后的进化守恒。前植物SCI。2016; 7:1800。
- 42。
亚当斯KL,温德尔JF。植物多倍体与基因组进化。植物生物学。2005;8(2):135–41.
- 43。
De Smet R,Van de Peer Y.基因组重复事件后遗传网络的冗余和重新定向。CurrOp植物BIOL。2012; 15(2):168-76。
- 44。
Moghe Gd,Shiu Sh。多倍体在开花植物中的原因及分子后果。Ann N Y ACAD SCI。2014; 1320:16-34。
- 45。
Ganko Ew,Meyers Bc,Vision TJ。拟南芥中复制基因之间的表达分歧。mol Biol Evol。2007; 24(10):2298-309。
- 46。
Lan T,Yang ZL,Yang X,Liu Yj,Wang XR,曾Qy。杨树谷胱甘肽S-转移酶超级家族的广泛功能多样化。植物细胞。2009; 21(12):3749-66。
- 47。
杨泽,刘海,王克,曾Qy。杨树聚乳糖醛酸糖酶超级家族棚灯对植物中越来越复杂的器官演化的分子演化和表达分歧。新植物。2013; 197(4):1353-65。
- 48。
Martinez M,Cambra I,Carrillo L,Diaz-Mendoza M,Diaz I.在大麦的整个胱抑素基因家族的表征及其靶线素L样半胱氨酸 - 蛋白酶,在种子萌发过程中Herdein动员的合作伙伴。植物理性。2009; 151(3):1531-45。
- 49。
Quain MD,Makgopa Me,Marquez-Garcia B,Comadira G,Fernandez-Garcia N,Olmos E等人。异位植物植物表达引起大豆(型)在大豆中增强的干旱胁迫耐受性甘氨酸最大)以及拟南芥蒂利亚纳通过对杂交途径的影响,也可以导致种子性状改善。植物Biotechnol J. 2014; 12(7):903-13。
- 50。
Arai S,Matsumoto I,Emori Y,Abe K。植物种子胱抑素及其内源和外源性靶酶。食品化学杂志。2002;50(22):6612–7.
- 51。
Carrillo L,Martinez M,Ramessar K,Cambra I,Castanera P,Ortego F等。大麦胱抑素基因在玉米中的表达通过改变其半胱氨酸蛋白酶增强对植食性螨的抗性。植物细胞代表2011;30(1):101–12.
- 52。
Martinez M,Lopez-Solanilla E,Rodriguez-Palenzuela P,Carbonero P,Diaz I.抑制大麦胱抑素HV-CPI(基因冰)抑制植物致病性真菌(基因冰)与其半胱氨酸蛋白酶抑制性能无关。mol植物微生物相互作用。2003; 16(10):876-83。
- 53。
波波维奇M,Andjelkovic U,Burzer L,Lindner B,Petersen A,Gavrovic-Jankulovic M.来自Green Kiwifruit的重组产生的抗真菌半胱氨酸蛋白酶抑制剂的生化和免疫表征(猕猴桃). 植物化学。2013;94:53–9.
- 54。
Shindo T,Kaschani F,Fan Y,KovácsJ,Fang T,Kourelis J等。非注释的假鼠霉素筛选筛选筛选筛选抑制番茄免疫蛋白酶的毒力因子。PLOS POAROG。2016; 12:E1005874。
- 55。
Gutierrez-Campos R,Torres-Acosta Ja,Sauceo-Arias LJ,Gomez-Lim Ma。使用半胱氨酸蛋白酶抑制剂在转基因烟草植物中对穴位的工程抵抗。自然生物科技。1999年; 17(12):1223-6。
- 56。
Okazaki S,Kaneko T,Sato S,Saeki K.通过Rhizobial III分泌系统劫持肉豆蔻瘤信号传导。《美国国家科学院学报》上。2013; 110:17131-6。
- 57。
Perez Guerra JC,Coussens G,De Keyser A,De Rycke R,De Bodt S,Van de Velde W等德尔。发育和应激诱导的结节衰老的比较截形苜蓿.植物理性。2010; 152(3):1574-84。
- 58。
Young ND、DebelléF、Oldroyd GE、Geurts R、Cannon SB、Udvardi MK等。Medicago基因组提供了根瘤菌共生体进化的见解。自然。2011;480(7378):520–4.
- 59。
李奇,张立,李,邓威尔·吉姆,张扬。比较基因组学表明,祖先的多倍体事件导致增强的根结节共生Papilionoideae..mol Biol Evol。2013; 30(12):2602-11。
- 60。
Chan SJ、San Segundo B、McCormick MB、Steiner DF。克隆的人和小鼠肝癌前体B cDNA的核苷酸和预测氨基酸序列。PNAS公司。1986;83(20):7721–5.
- 61。
Turk D,Guncar G.溶酶体半胱氨酸蛋白酶(组织蛋白酶):有前途的药物靶标。Acta CrystaPraphica D.生物晶体学。2003; 59(PT 2):203-13。
- 62。
刘兰,崔俊,张X,魏T,江普,王紫典。半胱氨酸蛋白酶的结构,功能和表位分析Spirometra erinaceieuropaei spargana..BioMed Res Int。2013; 2013年:198250。
- 63。
元,李河,王我,陈H,张c,陈l等。在大豆基因组中寻找结核和结节显影相关的胱抑素基因(甘氨酸最大)。前植物SCI。2016; 7:1595。
- 64。
Tamura K,SteCher G,Peterson D,Filipski A,Kumar S. Mega6:分子进化遗传学分析6.0版。mol Biol Evol。2013; 30(12):2725-9。
- 65。
Guindon S,Gascuel O.一种简单,快速,准确的算法,通过最大可能性来估计大的系统发育。系统中的生物学。2003; 52(5):696-704。
- 66。
ronquist f,teslenko m,van der mark p,Ayres DL,Darling A,Hohna S等人。MRBAYES 3.2:高效的贝叶斯系统发育推理和跨大型模型空间的模型选择。系统中的生物学。2012; 61(3):539-42。
- 67。
Stamatakis A. Raxml版本8:一种系统发育分析和大脑发育后的分析。生物信息学(牛津,英格兰)。2014; 30(9):1312-3。
- 68。
林奇M,科纳利JS。重复基因的进化命运和后果。科学(纽约州纽约市)。2000;290(5494):1151–5.
- 69。
杨忠。paml4:最大似然系统发育分析。分子生物学。2007;24(8):1586–91.
- 70
海勒曼J、莫蒂埃G、德帕佩A、斯佩尔曼F、范德桑佩尔J。qBase相对定量框架和软件,用于实时定量PCR数据的管理和自动分析。基因组生物学。2007;8(2):R19。
- 71
袁S,朱哼,苟哼,傅,刘l,陈t,等。结节子组织中涉及共生受体激酶的泛素连接酶。植物理性。2012; 160(1):106-17。
- 72。
冯毅,吴平,傅伟,彭立,朱赫,曹毅,等莲花japonicus.泛素连接酶Sie3与转录因子SiP1相互作用并形成同型二聚体。前植物SCI。2020; 11:795。
- 73。
陈磊,秦磊,周磊,李鑫,陈卓,孙磊,等。结节定位的磷酸盐转运体GmPT7在大豆中增强共生N(2)固定和产量方面发挥着重要作用。新植物。2019; 221(4):2013-25。
- 74。
Kereszt A,Li D,Indrasumunar A,Nguyen CD,NontachaiyApooms,Kinkema M等赛。根出杆菌根草杆菌介导的大豆转化研究根生物学。NAT PROTOC。2007; 2(4):948-52。
致谢
我们感谢廖红教授对RNAi载体p5941-35S-内含子(GFP-Bar-marker)的贡献。
资金
来自中国国家自然科学基金的资金(Grant No.11701346),中央非营利科学机构的基础资金(Grant No.1172018001),中国国家转基因项目(2016ZX08004-005)的基础研究资金(2016ZX08004-005),CAAS(CAAS-ASTIP-2016-OCRI)的农业科技创新计划,中国国家自然科学基金(Grant编号U1904102),南湖学者兴阳师范大学青年学者课程。资金机构在研究和收集,分析和解释方面没有在数据和写作稿件的设计中没有作用。
作者信息
隶属关系
贡献
S.Y.,D.K.,Y.G。和新安。Z.设计了这项工作,S.Y.写了稿件,S.Y.,D.K.,R.L.和X.L.执行大多数实验和分析,L.W.,H.C.,C.Z.,Y.H.,L.C.,Q.H.,H.Y.,D.C.,S.C.,W.G.,Z.S.,Z.Y.,Xiaojuan。Z.和D.Q.基本上贡献了这项工作的完成。所有作者都阅读并批准了最终手稿。
通讯作者
伦理宣言
道德认可和参与同意
不适用。
出版许可
不适用。
相互竞争的利益
提交人声明他们没有竞争利益。
附加信息
出版商的注意事项
斯普林格自然保持中立,就管辖权的要求,在出版的地图和机构的联系。
补充信息
附加文件1:表S1。
大豆的详细信息PLCPS..
附加文件2:表S2。
NIH / NLM / NCBI CD搜索工具的大豆PLCP的域分析。
附加文件3:表S3。
大豆42个GO基因的基因识别信息PLCPS..
附加文件4:表S4。
用于QPCR分析的QPCR分析选定的引物gmplcps..
附加文件5:表S5。
底漆glyma.04g190700- 特价RNAi分析。
附加文件6:图。S1。
5个大豆根瘤样品中4个参考基因的稳定性分析。
附加文件7:图S2。
28所选择的引物的熔化曲线gmplcps..
附加文件8:图S3。
筛选阳性转基因毛状根。
权利和权限
开放访问本文是根据知识共享署名4.0国际许可证授权的,该许可证允许以任何媒体或格式使用、共享、改编、分发和复制,只要您给予原作者和来源适当的信任,提供到知识共享许可证的链接,并指出是否进行了更改。本文中的图像或其他第三方材料包含在文章的知识共享许可证中,除非在材料的信用额度中另有说明。如果材料未包含在文章的知识共享许可证中,并且您的预期用途不受法律法规的允许或超出允许的用途,您将需要直接获得版权持有人的许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/by/4.0/.Creative Commons公共领域奉献豁免(http://creativecommons.org/publicdomain/zero/1.0/)适用于本文提供的数据,除非在数据的信用额度中另有说明。
关于这篇文章

引用这篇文章
袁,S.,柯,D.,李,R。等等。大豆木瓜样半胱氨酸蛋白酶的全基因组研究及其在根瘤共生中的表达分析。BMC植物BIOL.20,517(2020)。https://doi.org/10.1186/s12870-020-02725-5
已收到:
认可的:
出版:
关键词
- 基因组调查
- 瘤的发育和衰老
- 木瓜蛋白酶样半胱氨酸蛋白酶
- 根结节共生
- 大豆